|
This month has also seen the publication of another paper of mine (in collaboration with Leigh Simmons), again in Proceedings B, but this time using micro-CT to examine the timing and location of female copulatory wounding in Callosobruchus maculatus. For this project I performed micro-CT scans of 19 mating pairs of C. maculatus that had been flash-frozen in liquid Nitrogen at different stages of mating. We then used these scans to examine the interactions between male and female genitalia during mating, and visualise these interactions in unprecedented detail. Importantly, we were able to use the scan data to detect copulatory wounding of females without having to wait for an immune response, which is needed when assessing wounding using a light microscope. We could therefore use the micro-CT technique to examine how tract damage accumulates during mating, and how the timing of damage relates to female mating behaviour. This paper is also a showcase for the power of micro-CT to aid our understanding of reproductive behaviour and morphology, and I hope it inspires more people to use this technique in the future! Please follow this link, or the relevant link on the publications page, to read the full paper.  Example figure from the paper, showing the location of female copulatory wounding (circles) in relation to the male penis spines, across 19 mating pairs.
0 Comments
I'm very excited to say that my newest paper was published online a few weeks ago in Proceedings B, looking at sexual conflict in the seed beetle Callosobruchus maculatus. This is the first paper from my work at UWA, and is the result of a collaboration with several researchers at UWA (Emile van Lieshout, Kathryn McNamara & Leigh Simmons) as well as Göran Arnqvist at Uppsala University. The project benefitted especially from the use of C. maculatus lines that Göran has been maintaining at Uppsala for many years. In the paper we show that there has been correlated evolution between the size of the male penis spines and three defensive female adaptations (the thickness of the female reproductive tract lining, and two measures of female immune activity), across 13 C. maculatus population that have been isolated in the lab for over 100 generations. Importantly, we also show that the amount of copulatory wounding females receive during mating is related to the relative thickness of the female reproductive tract in relation to male spine length (as shown in the image below). This is strong evidence that males and females of this species are locked in an evolutionary arms race driven by sexual conflict. Follow the link here to read the paper, or go to the publications page for the pdf. 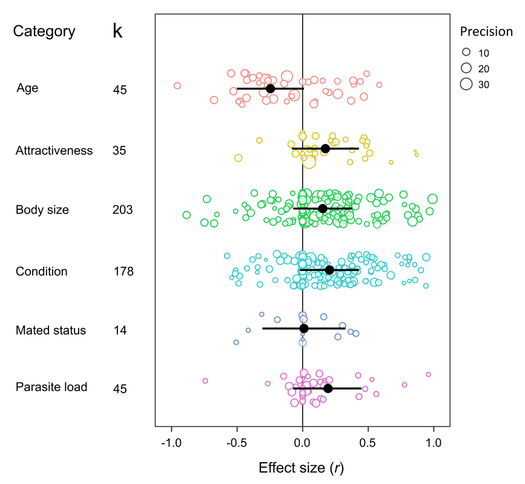 A few months ago I wrote here about my work using X-Ray micro-CT to study the coevolution of male and female genitalia in insects. I also used that post to present some early images of the male and female genitalia of the seed beetle Callosobruchus maculatus I made from those scans. In this post I would like to present some more CT images, but focusing on a different species: the Indian Meal Moth Plodia interpunctella. This time we are looking at the interaction between the region of the female reproductive tract where the male deposits his sperm (known as the bursa), and the structure containing the male’s sperm, known as the spermatophore. In several insect groups, including butterflies and moths, the male sperm is not deposited freely but is instead transferred in a discrete package known as a spermatophore. In Plodia interpunctella the spermatophore has an elaborate structure. Below is a 3d volume rendering of a spermatophore inside a female. The spermatophore has two main parts: a hollow, spherical sac which contains the sperm mass, and an elaborate chitinous neck.  3d reconstruction (false-colour) of a male spermatophore inside the female bursa after mating. Note the spherical sac (right) and long neck region ending with a hole (left). The spermatophore is passed to the female in liquid form, which then solidifies and hardens during mating into the characteristic shape seen here. Inside the spherical sac are around 30 large sperm bundles, each formed of many sperm cells (see below). Once in the spermatophore has been deposited, the sperm move along the neck and out of the hole at the top.  3d reconstruction of the sperm bundles tightly packed inside the spermatophore. Once the sperm has been transferred to the female the spermatophore shrinks but is not broken down by the female (see below). This is useful because it means that we can count the number of times a female has mated (either in the wild or in the lab) by dissecting out the bursa after she has died and counting the number of spermatophores present. 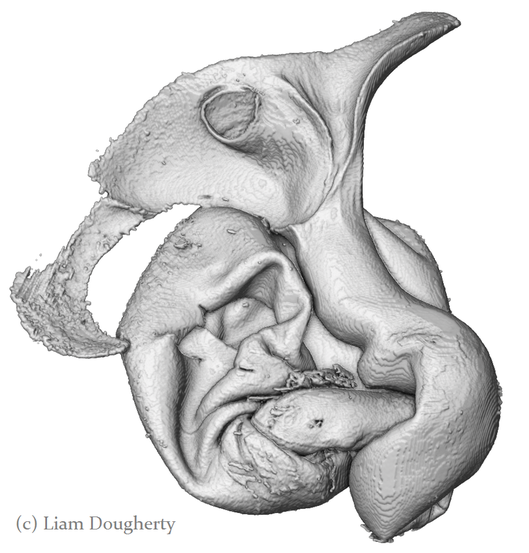 3d reconstruction of an old, shrunken spermatophore inside the bursa of a female. These can be counted to determine the number of times a female has mated. The female bursa is a large hollow organ into which the spermatophore is deposited. Sperm are not stored here, but instead are stored in a much smaller organ called the spermatheca. To get to the spermatheca the sperm need to be moved along a narrow duct which opens into the bursa. Surrounding the entrance to this duct on the wall of the bursa is a row or sharp teeth known as lamina dentata or signa (see below). The teeth vary in size and shape within this species.  3d reconstruction of the female bursa, with a virtual slice to show the internal structure. Note the row of teeth next to the spermathecal duct entrance. In other moth and butterfly species these teeth are used to physically pierce the spermatophore capsule, thus releasing the spermatophore contents into the bursa. One reason why females may want to pierce the spermatophore is because the presence of a full spermatophore in the bursa may prevent the female from remating. Males generally benefit from preventing female remating as it may reduce his paternity share. Therefore one hypothesis fur the function of bursal teeth is that they allow the female to regain some control of her remating rate by speeding up the breakdown of the spermatophore. Males could then evolve larger or thicker spermatophores in response, leading to an ‘arms race’ between bursal anatomy and spermatophore anatomy. I have been using micro-CT to examine potential coevolution between bursal teeth morphology and spermatophore morphology in the Indian Meal Moth. This work is being done in collaboration with Leigh Simmons at Kathryn McNamara at UWA, and Nina Wedell at the University of Exeter. Watch this space for news on future publications! During the last year I have been working at UWA on the use of Micro-CT X-Ray scanning for studying the coevolution of insect genitalia. Micro-CT is not yet widely used in the field of genital evolution and we are hoping to be able to showcase the types of analysis that are possible when using modern scanning hardware and visualisation software. Micro-CT has several advantages over traditional anatomical techniques. First and foremost, it allows for the creation of three-dimensional datasets that can be shared online and viewed (and manipulated) by anyone using free open-source software. A key advantage for us is the ability to visualise internal anatomical structures without dissection, which runs the risk of damaging or altering the shape of such structures. In this post I would like to present a few examples of the kinds of visualisations that I have been doing, using as an example our work on the seed beetle Callosobruchus maculatus. Below is a visualisation of a pair of C. maculatus scanned during mating. The male is on the left and the female on the right. To create this image, a mating pair were first flash-frozen in liquid Nitrogen, so as to freeze the genitalia in place. The pair were then placed into a staining solution in order to increase the X-Ray contrast of the tissues, making analysis much easier. I have virtually removed the lowest density tissues, which has the result of making the cuticle transparent. The tissue has also been colour-coded by density, with highest density tissue in red and lowest in blue. The male leg muscles can be seen in red/yellow (muscle tissue is not very dense but takes up stain very well, resulting in high X-Ray absorbance). The large spherical structure in the female abdomen is the reproductive tract, which has expanded as the male has already deposited his spermatophore. 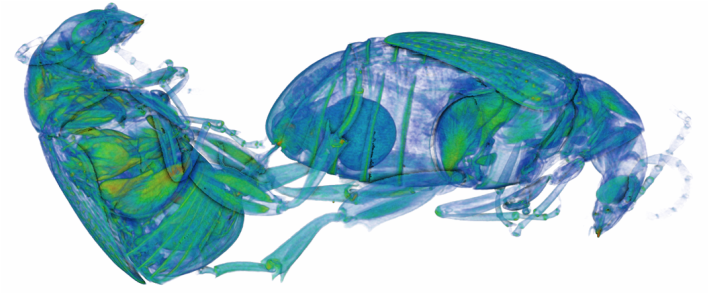 C. maculatus mating pair C. maculatus males are probably best known for their penis, which is covered in sharp spines. Below is a visualisation of a C. maculatus penis that has been ‘virtually dissected’ from the female reproductive tract during mating. The rest of the male is to the left, and the head of the penis is to the right (sperm is released from here). Again this has been colour-coded by density. The spines can be seen in red, meaning they spines are made of a high density material when compared to the rest of the penis. Note also the number of spines; I counted around 230 on this male (it is hard to be exact because some spines are very small). 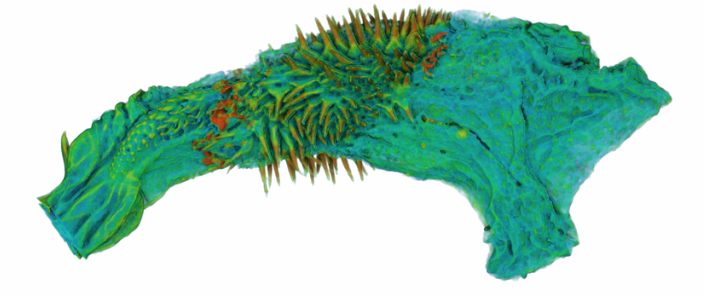 C. maculatus penis inside female During mating the spines on the penis stab into the wall of the female reproductive tract, leaving deep scars and wounds. This is harmful to the female, and can result in a significant reduction in female fecundity and even lifespan. Consequently, in this species the walls of the female reproductive tract are thickened when compared to closely related species in which the males lack penis spines. Below is a visualisation of the area of the female reproductive tract contacted by the penis spines during mating, which I have virtually sliced length-wise. The end of the female abdomen is to the left and the female head is to the right. The wall of the tract is very thick here when compared with the regions that are not in contact with the penis spines. The dark line in the centre is the lumen of the tract, which is very narrow when not mating (this is expanded greatly when the penis enters). 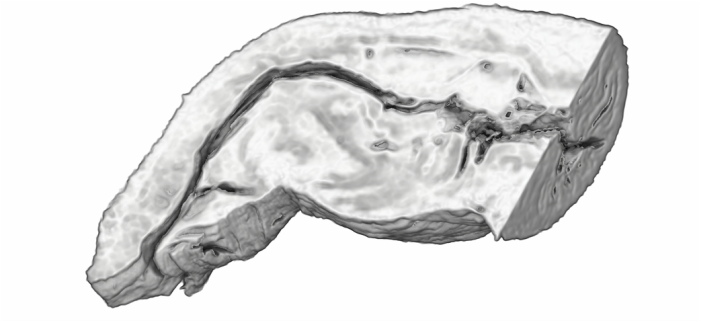 Reproductive tract of female C. maculatus I have been using micro-CT to measure changes in the thickness of the female reproductive tract in C. maculatus across populations and in response to different experimental conditions. Micro-CT has allowed us to do things previously not possible using dissection, such as calculating the volume of tissue in a given morphological trait. I have also been scanning pairs in copula to look at the interactions between male and female genitalia during mating, and the timing and location of female tract scarring. This is all still a work in progress but hopefully it will begin to be published soon, along with plenty of high resolution images and videos. For now watch this space for more info! NOTE: all of the above scans were performed using a ZEISS Versa 520 XRM at the University of Western Australia, and visualised using the AVIZO software package. All samples were stained with iodine prior to scanning. The voxel size is 7.4 μm3 for the mating pair, 1.45μm3 for the penis and 2.35 μm3 for the female tract. The recent commentary piece I wrote in collaboration with Leigh Simmons and Dave Shuker has just been published online in Animal Behaviour. In it we make the point that not all females in a population have to mate more than once for post-copulatory sexual selection to occur. You can read the full text here (open access for the next 50 days) or on the publications page.
My recent paper (with Dave Shuker) in Behavioral Ecology and Sociobiology is now online. In it we measure the strength of selection on the length of the penis in two lygaeid bug species. Interestingly, in one of the species we found pre-copulatory selection on penis length, which was unexpected given that the penis is stored inside the body prior to mating. The strength of this selection was also influenced by the number of males competing for each female. You can read it the full-text here or here.  A meta-analysis I was involved in has just come out online in Behavioral Ecology this week. This one was led by Fhionna Moore from the University of Dundee, with a bit of help from me and Dave along the way. In it we tested whether physiological stress influences the strength of sexual signals across a range of vertebrate species, and show surprisingly that no, the relationship is basically zero. You can read all about it here. The paper has been accepted as an invited review, which means it is open access. There will also be several invited commentaries published online in the near future so look out for those.  Our paper on penis breakage in Lygaeus simulans was published online this week in the European Journal of Entomology. In it we show that almost a quarter of males that were able to mate often have broken penises, and breakage probably occurs after mating not during. However it may not be all bad for these males; find out why by reading the paper here.  Last month I arrived in Perth, Australia to start a postdoc with Leigh Simmons at The University of Western Australia. I plan to look at the evolution of female reproductive tract morphology in the seed beetle Callosobruchus maculatus and the Indian meal moth Plodia Interpunctella.  A view of the centre of Perth from the UWA campus. Last week I was in Lausanne, Switzerland for the European Society of Evolutionary Biology's biennial conference. It was a tiring but super-interesting five days, during which I saw some great talks. Here is me standing next to the penis-snipping poster that I brought along (courtesy of Paris Veltsos). Between the science, the conference bar and the great Swiss food (lots of cheese!) it was a great week!  |
Archives
February 2024
AuthorLiam Dougherty. Categories |
 RSS Feed
RSS Feed
