|
Our new paper is now online in Ecological Solutions and Evidence. We produce a systematic map of studies testing the link between environmental temperature and some form of animal reproduction. We find over 1,600 studies, but lots of research gaps. This was the result of a great collaboration between members of the European Thermal Fertiltity Network. We hope to use this databse for several projects- watch this space! 
0 Comments
I am now considering applicants for a fully-funded PhD vacancy starting in September 2024. The project is in collaboration with Tom Price and David Atkinson at Liverpool, and Nicola Hemmings at Sheffield.
, examining the male fertility gap in Drosophila fruit flies. The application deadline is January the 8th 2024, and is open to both UK and non-UK applicants. See the link for more details on the application process, and please contact me if you have any questions about the project itself. New paper: impact of heat stress on male courtship behaviour and mate choice in Drosophila virilis8/9/2023 This week saw the publication of Kwan's first paper from her master's research, supervised by Tom Price and I. Kwan looked at the effect of heat stress on male mating behaviour in the Asian fruit fly species Drosophila virilis. She found that heat-sterilised males don't change their courtship behaviour, but do have reduced mating success. This suggess that females can tell something about male fertility, though we couldn't determine how. Nevertheless, female mate choice might mitigate the population-level cost of heat stress in this species. Please click through to read more! 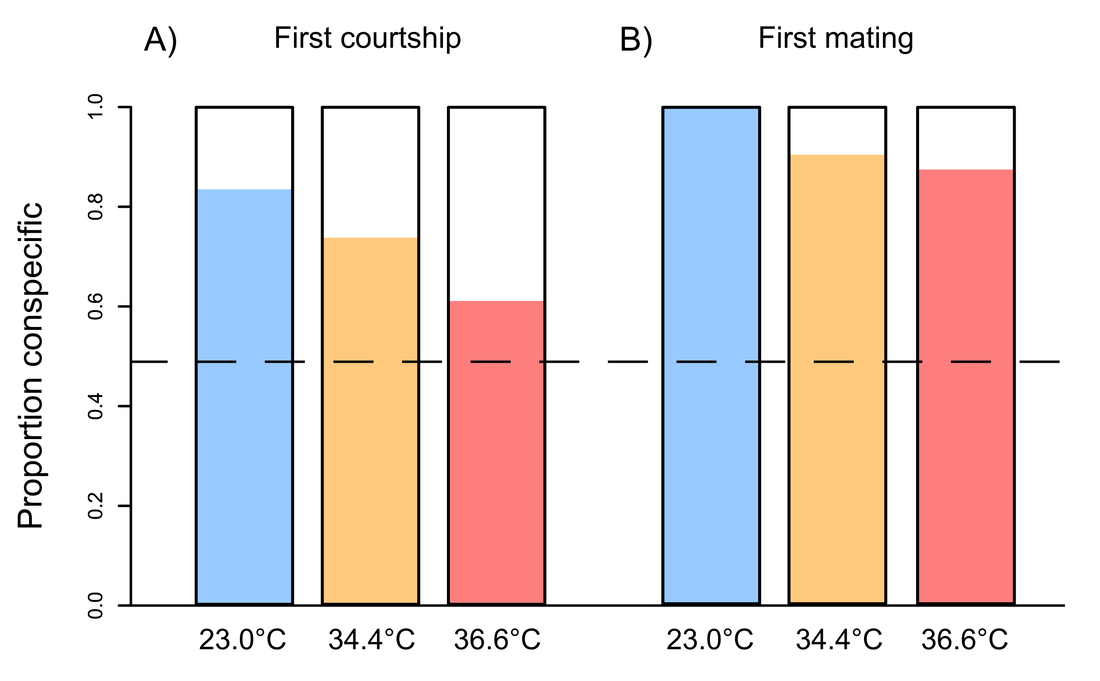 Our new paper was published online this week. We present a meta-analysis quantifying the correlation between parasite load and the expression of sexual ornaments across the animal kingdom. Interestingly, we find a significant negative relationship, but only for parasites that are able to transmit during sex, or for ornaments that can be expressed dynamically. This was a great collaboration led by Jess Stephenson at Pittsburgh 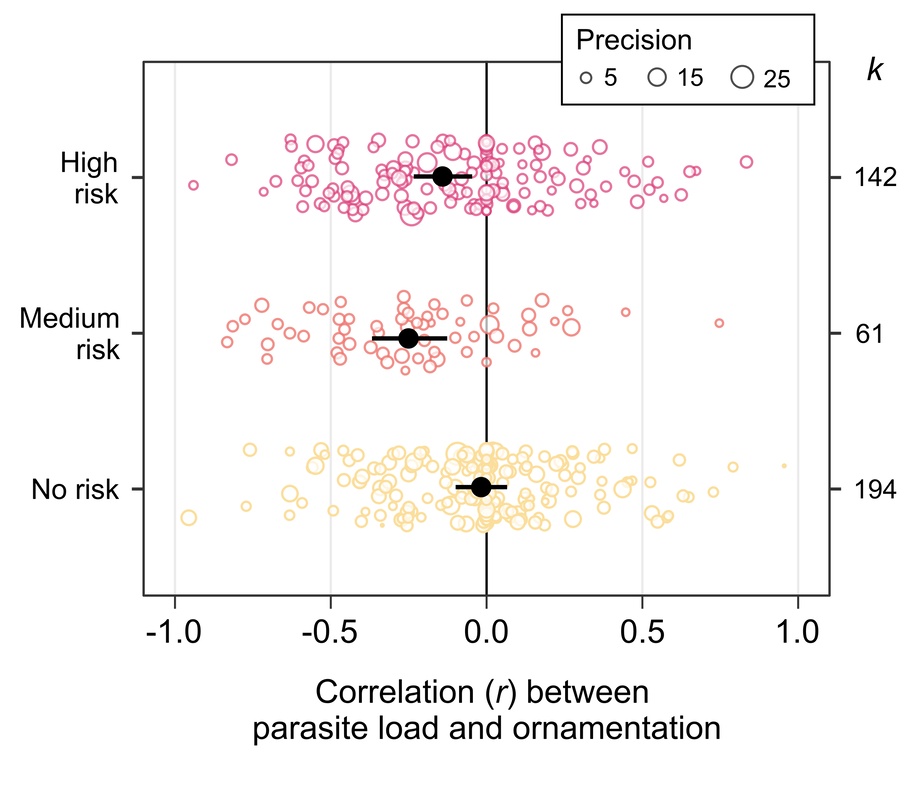 I am a co-supervisor on this project focusing on how the environment drives the evolution of plumage and other sexual traits in birds. This project is led by Dr Chris Cooney, and is based at the University of Sheffield. Current application date is May 8th, and is only eligible for UK-based applicants. See the link for more details.
This week Fay Frost joined the lab as a post-doc on a NERC-funded project focusing on using meta-analysis to quantify the reduction in animal fertility following exposure to high temperatures. Welcome Fay!
This week saw the publication of my latest meta-analysis in Behavioral Ecology. I summarise the evidence that individual state influences the strength of mate choice in animals. Overall, the evidence for state-dependent mate choice is stronger for females compared to males, though the mean correlation is similar for both sexes. See the link above, or the pdf on my publications page, to read more.  I will soon advertise a 9-month postdoc position, working in my lab in collaboration with Dr Tom Price. In the role you will investigate how high temperatures affect male and female fertility across the animal kingdom, by analysing a large data set of published papers collected in the last year. Hybrid and remote working arrangements will be considered.
The post will be advertised in January, and the latest start date is April 1st. So please contact me now if interested! I am a co-supervisor on three funded PhD projects being advertised for September 2023. All are funded through the NERC ACCE doctoral training scheme, and have the same application deadline: the 13th January 2023.
The first project looks at how the environment influences male sperm allocation strategies in fruit flies. The project is led by Dr Stuart Wigby, and the student would be based at the University of Liverpool. The second project concerns how temperature extremes influence mosquito fertility and virus transmission in the UK. This project is led by Dr Marcus Blagrove, and is also based at the University of Liverpool. The third project concerns how the environment drives the evolution of plumage and other sexual traits in birds. This project is led by Dr Chris Cooney, and is based at the University of Sheffield. Please see the links for details, or here for all three: https://www.findaphd.com/phds/?Keywords=dougherty And feel free to contact me for more details. |
Archives
February 2024
AuthorLiam Dougherty. Categories |
 RSS Feed
RSS Feed
